
几乎所有动物都不可避免死亡的结局,生命的长度为何是有限的?为什么有些动物寿命长而有些动物寿命短?本篇文章将通过对于长寿动物的基因进行分析,从而从基因层面上去解释动物寿命有长短区别的原因。
哺乳动物的寿命存在一个规律:体重越大,寿命越长,原因可能是跟基础代谢率有关。这里有一张说明了体重和寿命相关性的图:
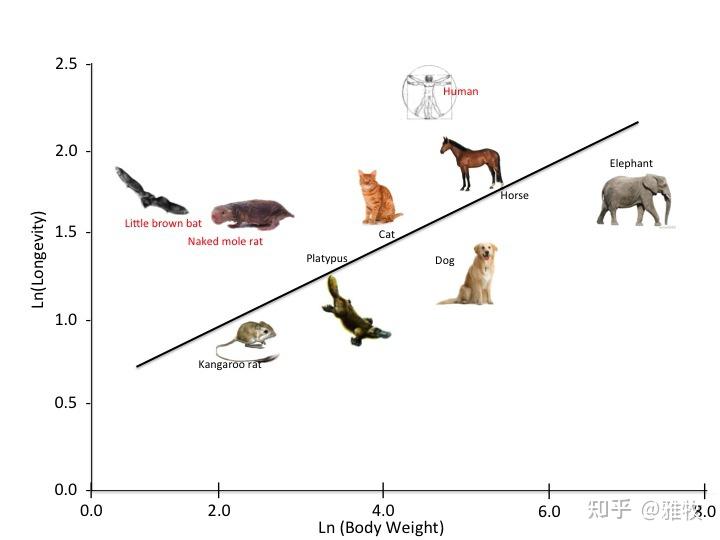
图片来源:The bat blueprint - On Biology (biomedcentral.com)
如图所示,人类、裸鼹鼠、小棕蝙蝠的相对寿命是比较长的。
布氏鼠耳蝙蝠
新冠肺炎的流行和新冠肺炎病毒来源于蝙蝠的猜测引起了人们对于蝙蝠这类动物的关注,蝙蝠具有强大的免疫能力,这使得他们可以和各种病毒共存,蝙蝠还有一个神奇的现象——普遍长寿,其中,鼠耳蝙蝠(属)是其中的佼佼者,而鼠耳蝙蝠中的布氏鼠耳蝙蝠(Myotis brandti) 寿命竟然长达40年以上(目前已经发现寿命最长的布氏鼠耳蝙蝠是41岁,而且这只蝙蝠是野生的)并且很少得癌症,我们知道,体型小的哺乳动物寿命往往较体型大的哺乳动物短命,布氏鼠耳蝙蝠非常小,仅有 33~37 mm长,体重仅有7~8g,实验室常用的小鼠体重20g左右,比布氏鼠耳蝙蝠还大点,寿命却仅有2~3年,已知哺乳动物中,只有19种相对(考虑到体型因素)人类来说有更长的寿命,其中包括裸鼹鼠和18种蝙蝠,而布氏鼠耳蝙蝠是已知哺乳动物中,相对其体重而言寿命最长的。
鼠耳蝙蝠如此长寿的原因是什么?针对鼠耳蝙蝠的基因研究为我们揭示了鼠耳蝙蝠长寿的机制,我把已知鼠耳蝙蝠长寿的主要机制,分成四个方面来说:
1、IGF-1受体和生长激素受体基因突变:
研究发现,布氏鼠耳蝙蝠的IGF-1受体和生长激素受体基因突变导致功能障碍,同样的突变若出现在人类身上将导致侏儒症,这可以解释布氏鼠耳蝙蝠体型为何如此微小。针对人类侏儒症患者的研究显示他们更不易得糖尿病和癌症,针对秀丽隐杆线虫的研究发现,携带DAF-2/IGF-1受体以及TOR效应蛋白RKS-1/S6K的功能缺失突变的秀丽隐杆线虫能够大幅度延长寿命(达到五倍之长),IGF-1受体和生长激素受体功能障碍,或许是布氏鼠耳蝙蝠长寿的一个重要原因。
2、强大的DNA修复能力:
研究发现,鼠耳蝙蝠们具有更高的ATM和SETX表达,这两个基因涉及基因修复能力。ATM是直接感受DNA双链断裂损伤并起始诸多DNA损伤信号反应通路的主开关分子,ATM基因的突变会导致运动失调性毛细血管扩张症(ataxia telangiectasia)并增加癌症风险。之前有研究发现,增强ATM活性可以延缓早衰模型小鼠衰老并延长其寿命。而SETX错义突变会导致肌萎缩侧索硬化(amyotrophic lateral sclerosis, ALS)。
3、强大的维持端粒长度的能力:
我们知道,包括人类在内的绝大多数哺乳动物的端粒随着细胞的分裂而缩短,随着年纪增长,我们的端粒越来越短,而根据鼠耳蝙蝠端粒长度和年龄之间相关性的研究,鼠耳蝙蝠端粒长度并不随着年龄的增长而下降,研究发现,鼠耳蝙蝠并没有表现出更高的端粒酶活性,但是ATM、MRE11a、RAD50 和 WRN这几个和“端粒延长替代机制(alternative lengthening of telomeres,ALT)”相关的基因表达水平比较高。对于绝大多数哺乳动物而言,仅在一些异常情况,比如癌细胞中才会出现ALT机制,而鼠耳蝙蝠似乎在正常情况下就能启动ALT机制从而使自己的端粒不缩短。
4、抗衰老相关miRNA的差异表达:
针对鼠耳蝙蝠血液里的miRNA研究发现,鼠耳蝙蝠的一些与衰老、癌症相关的miRNA表达与其他哺乳动物不同,比如说,鼠耳蝙蝠高表达具有肿瘤抑制作用的miR-101、miR-16 和miR-143,并且高表达能够上调NRF2功能的miR-155,而NRF2具有抗氧化应激的作用。
裸鼹鼠
裸鼹鼠(Heterocephalus glaber)是另一种以长寿著名的哺乳动物。这种丑陋的动物体型和大鼠差不多大,但其长达三十年的寿命是大鼠的十倍,它们衰老速度缓慢,且很少得癌症,这使得它们成为了衰老领域的热门研究对象,那么裸鼹鼠具有哪些优势基因呢?研究发现:
1、端粒相关基因优势
裸鼹鼠身上存在TOP2A、TEP1 和 TERF1 三个与端粒相关基因的正向选择优势,这三个基因属于端粒路径交替延长的五种蛋白复合物相关基因的一部分。亦有研究发现,裸鼹鼠的TRF1基因(端粒重复序列结合蛋白质1基因)存在一个特殊的氨基酸变异,这使得裸鼹鼠的TRF1蛋白与端粒的结合能力更强并更好得维护端粒,已知TRF1随着年龄增长而降低,过表达TRF1可以延缓衰老,TRF1基因的突变可能是裸鼹鼠长寿的一个因素。有趣的是,裸鼹鼠的TERT(端粒酶逆转录酶)基因可以随着年龄增长依然稳定表达。多项研究显示,裸鼹鼠的端粒相对稳定,不会随着年龄的增长而缩短。
2、有利于长寿的基因表达变化趋势
研究发现,人身上的一些基因表达随着年龄的增长而发生变化,而裸鼹鼠身上的相应基因表达则比较稳定,有些基因变化则是相反的,比如说在人身上,CYP46A1和SMAD3这两个基因随着年龄增长而降低,在裸鼹鼠身上却随着年龄增长而上涨了。CYP46A1基因的产物是胆固醇内稳态的介质,它影响胆固醇的变化趋势β 聚集。SMAD3的产物是TGF-β信号的调节器,通过减缓细胞增殖速度在癌症发展中发挥作用。衰老过程中裸鼹鼠体内的SMAD3的高表达可能有助于优化细胞死亡率,保护裸鼹鼠免受癌症的侵袭。与大分子降解相关的基因,比如GSTA1、DERL1 和GNS,在裸鼹鼠身上不随着年龄增长而上涨。NDUFB11、 ATP5G3和 UQCRQ等与线粒体蛋白相关基因不随着年龄增长而下降,这可能有助于维持裸鼹鼠的线粒体功能。
3、更强的DNA修复能力与相关基因
一项针对裸鼹鼠和小鼠的DNA修复能力的研究显示,裸鼹鼠有更有效率的切补修复能力。另一项针对18种啮齿动物的DNA修复能力的研究显示,裸鼹鼠和河狸等相对长寿的啮齿动物有更强的DNA修复能力,它们的DNA修复能力与SIRT6活性有关,并且人类的SIRT6活性比小鼠等短命动物要强一些。
4、强大的抗癌能力与相关基因
前面我们提到,裸鼹鼠的癌症相关基因表达有利于抗癌,另有研究发现,2020年,一项研究发现裸鼹鼠胚胎成纤维细胞(NEFs)中p53蛋白的半衰期是小鼠和人胚胎成纤维细胞中p53蛋白半衰期的10倍以上,这项研究表明,p53蛋白的独特稳定和调节可能有助于裸鼠对癌症的显著抵抗。这篇研究亦提到,系统发育分析表明,裸鼹鼠的基因组中含有17个磷酸酯酶和张力蛋白(PTEN)假基因拷贝,这可能是裸鼹鼠优秀抗癌能力的原因之一。
弓头鲸
弓头鲸(Balaena mysticetus)是一种长寿的大型海洋哺乳动物,它们拥有巨大的头部和造型独特的弓形下颚,它们生活在北半球的寒冷水域,它们的寿命可超过两百年,是已知寿命最长的哺乳动物。弓头鲸如此长寿,它们具有哪些方面的基因优势呢?
一项针对弓头鲸和小须鲸的比较基因组分析之正选择分析(dN/dS)研究显示,弓头鲸在以下基因方面具有正选择优势:
细胞因子信号传导抑制因子2(SOCS2)、抑制素(APTX)、头蛋白(NOG)和瘦素(LEP)。和尖嘴海豚相比,正选择价值前5%的基因包括:
叉头盒状O转录因子(FOXO3)基因、切割修复交叉互补3(ERCC3)基因和成纤维细胞生长因子受体1(FGFR1)基因。
细胞因子信号转导抑制因子是SOCS家族中的一员,参与调节包括个体生长、代谢、骨形成、肿瘤发生、机体免疫在内的许多生理过程有关。FOXO3是著名的长寿基因,当机体处于氧化应激态时,FOXO3 会受到修饰从而入细胞核,启动氧化应激反应相关基因的表达,从而保护细胞免受压力损伤。切割修复交叉互补(ERCC)基因家族则在DNA自我修复过程中起着关键作用。
为了找出弓头鲸蛋白特有的氨基酸变异,这项研究在弓头鲸和其他九种哺乳动物之间做了同源序列对比,以这种方式鉴定的谱系特异性残基先前已被证明是蛋白质功能显著变化的指示,这项分析揭示了几种与衰老和癌症相关的蛋白质,其中包括切割修复交叉互补1(ERCC1)、组蛋白去乙酰化酶1(HDAC1)、组蛋白去乙酰化酶2(HDAC2),前面我们已经提到切割修复交叉互补(ERCC)基因家族的作用,而组蛋白去乙酰化酶在染色质结构和转录调控中起重要作用。
2025 补充:
2024年的一篇新研究显示,与其他哺乳动物相比,弓头鲸细胞修复双链断裂(DSB)和错配的效率和准确性更高。与保护免受基因毒性应激有关的CIRBP蛋白在弓头鲸体内的含量比其他哺乳动物高得多。并且,CIRBP及其下游蛋白RPA2(在弓头鲸细胞中也高水平存在)能够提高人类细胞DNA修复的效率和保真度。
象龟
加拉帕戈斯象龟是寿命最长的脊椎动物之一,这类庞大且笨重的陆龟的寿命可超过一百岁。2012年,一只叫孤独的乔治的加拉帕戈斯象龟去世了,自1971年被发现到2012年确认死亡为止,被认为是平塔岛象龟(Geochelone nigra abingdoni)中已知的最后一个个体,孤独的乔治已经死亡,标志着平塔岛象龟至此绝种。
2019年一项针对孤独的乔治的基因的研究发表了,这项研究,用了孤独的乔治和另一个亚达布拉象龟(Aldabrachelys gigantea)的基因样本与其他物种进行对比,发现了一系列影响DNA修复基因、炎症介质和癌症发生相关基因的谱系特异性变体。具体包括:
DNA维护相关基因:GAPDH基因、NEIL1基因、RMI2基因和XRCC6基因等。
肿瘤抑制相关基因:SMAD4基因、NF2基因、PML基因、 PTPN11基因 和 P2RY8基因等。
端粒维持相关基因:DCLRE1B基因和TERF2基因等。
其他抗衰老基因:EEF1A1基因等。
百岁老人
值得一提的是,2021年5月发布的一项针对百岁老人的全基因组测序分析研究发现,寿命超过105岁的长寿老人和普通老人相比,百岁老人的基因让他们有更强的DNA修复能力、对抗活性氧的能力与更好的线粒体功能。
综合分析
根据以上针对不同长寿动物的研究,我们很容易发现,长寿动物普遍具有以下共性:
强大的端粒维持功能、强大的DNA修复能力、强大的抗癌能力。为什么这三大因素与寿命有关?我们看看这三大因素是如何影响寿命的:
端粒是真核细胞染色体末端的特殊结构,细胞每分裂一次,端粒就缩短一点,当端粒缩短到一定程度,细胞将失去分裂能力,维持端粒完整性对于细胞生存非常重要,所以想要活得长,端粒维持能力就要强,这或许可以解释为什么长寿动物的端粒相关基因普遍具有正向选择优势。随着动物的发育和成长,因为各种内因(线粒体产生活性氧自由基对DNA造成损伤)和外因(各种有毒分子进入体内破坏DNA),DNA面临损伤的风险,而DNA突变会引起端粒缩短、细胞衰老、凋亡、癌变等各种问题,因此对于动物维持健康身体而言,DNA修复能力至关重要。动物若患有癌症,很快就会去世,因此想要活得长,抗癌能力就得强。
长寿基因的优势是在长期的演化过程中缓慢形成的,在地球这颗蔚蓝色的星球上,从来没有哪个物种像人类这样知晓基因的秘密,在如今这个时代,我们已经初步掌握调控基因的技术,近年来基因治疗技术正在快速发展,尤其是新冠肺炎的流行让核酸药物走进了大众和投资人的视野,在不久的将来,我们或许可以通过调控基因从而使我们变得更加健康和长寿。
参考文献:
[1] Lan J , Rollins J , Wu D , et al. Translational regulation of non-autonomous mitochondrial stress response promotes longevity[J]. Cell Reports, 2019, 28(4):1050-1062.e6.
[2] Seim I , Fang X , Xiong Z , et al. Genome analysis reveals insights into physiology and longevity of the Brandt's bat Myotis brandtii[J]. Nature Communications, 2013, 4:2212-2212.
[3] Qian M , Liu Z , Peng L , et al. Boosting ATM activity alleviates ageing and extends lifespan in a mouse model of progeria[J]. eLife Sciences, 2018, 7:e34836-.
[4] Huang Z , Jebb D , Teeling E C . Blood miRNomes and transcriptomes reveal novel longevity mechanisms in the long-lived bat, Myotis myotis[J]. Bmc Genomics, 2016, 17(1):906.
[5] Foley N M , Hughes G M , Huang Z , et al. Growing old, yet staying young: The role of telomeres in bats' exceptional longevity[J]. Science Advances, 2018, 4(2):eaao0926.
[6] Kim E B , Fang X , Fushan A A , et al. Genome sequencing reveals insights into physiology and longevity of the naked mole rat.
[7] Augereau A , Mariotti M , Pousse M , et al. Naked mole rat TRF1 safeguards glycolytic capacity and telomere replication under low oxygen[J]. Science Advances, 2021, 7(8):eabe0174.
[8] Evdokimov A , Kutuzov M , Petruseva I , et al. Naked mole rat cells display more efficient excision repair than mouse cells[J]. Aging, 2018, 10(6):1454-1473.
[9] Tian X , D Firsanov, Zhang Z , et al. SIRT6 Is Responsible for More Efficient DNA Double-Strand Break Repair in Long-Lived Species[J]. Cell, 2019, 177(3):622-638.e22.
[10] Leonida S , Bennett N C , Leitch A R , et al. Patterns of telomere length with age in African mole-rats: New insights from quantitative fluorescence in situ hybridisation (qFISH)[J]. PeerJ, 2020, 8(13):e10498.
[11] Deuker M M , Lewis K N , Ingaramo M , et al. Unprovoked Stabilization and Nuclear Accumulation of the Naked Mole-Rat p53 Protein[J]. Scientific Reports, 2020, 10(1).
[12] Keane M , Semeiks J , Webb A E , et al. Insights into the Evolution of Longevity from the Bowhead Whale Genome[J]. Cell Reports, 2015, 10(1):112-122.
[13] Firsanov D, Zacher M, Tian X, Sformo TL, Zhao Y, Tombline G, Lu JY, Zheng Z, Perelli L, Gurreri E, Zhang L, Guo J, Korotkov A, Volobaev V, Biashad SA, Zhang Z, Heid J, Maslov A, Sun S, Wu Z, Gigas J, Hillpot E, Martinez J, Lee M, Williams A, Gilman A, Hamilton N, Haseljic E, Patel A, Straight M, Miller N, Ablaeva J, Tam LM, Couderc C, Hoopman M, Moritz R, Fujii S, Hayman DJ, Liu H, Cai Y, Leung AKL, Simons MJP, Zhang Z, Nelson CB, Abegglen LM, Schiffman JD, Gladyshev VN, Modesti M, Genovese G, Vijg J, Seluanov A, Gorbunova V. DNA repair and anti-cancer mechanisms in the long-lived bowhead whale. bioRxiv [Preprint]. 2024 Nov 5:2023.05.07.539748. doi: 10.1101/2023.05.07.539748. PMID: 39574710; PMCID: PMC11580846.
[14] Quesada V , S Freitas-Rodríguez, Miller J , et al. Giant tortoise genomes provide insights into longevity and age-related disease[J]. Nature Ecology & Evolution.
[15] Garagnani P , Marquis J , Delledonne M , et al. Whole-genome sequencing analysis of semi-supercentenarians[J]. eLife Sciences, 2021, 10:e57849.